






September 29, 2016
If you’ve been following our blog, you know that we recently initiated the hosting of a series of blogs, with permission of the author, from the website Vaccine Papers that compile the scientific proof that vaccines can and do cause autism. If you recall, Part 1 was about how immune activation (whether in pregnant mothers or in infants during the postnatal period when the brain and neurological system are in a phase of rapid development) can cause autistic behaviors and specific brain changes that have been shown to be characteristic of autism. Part 2 covered the mouse studies that make it clear that the presence of one specific cytokine, interleukin-6 (IL-6) in high levels in the brain (which is the case for autistic human brains) is a necessary and sufficient condition for the development of autism in mice. Part 3 was about the supportive studies done on primates.
Since many of the animal studies cited in Parts 2 and 3 specifically involved maternal immune activation during the prenatal period, in order to demonstrate that vaccinating children can cause autism it is important to establish that not only can such high levels of cytokines be generated postnatally (during human infancy and toddlerhood), but that those cytokines can disrupt brain development in a similar manner as well. Part 4 presents a wide range of scientific evidence that makes it clear that postnatal immune activation can indeed cause high levels of cytokines in the brain which can disrupt brain development in ways similar to that of maternal immune activation. The original is located here.
Those who can make you believe absurdities, can make you commit atrocities.
–Voltaire, French enlightenment philosopher and author, 1765. . . increased inflammatory state during early postnatal weeks is responsible for altered synaptic connectivity and impaired behavior . . .
–Coiro et al., 2015, in study cited below
Vaccine advocates, aware of the research on immune activation, have argued there are some obvious differences between the maternal immune activation (MIA) discussed in previous articles and vaccines:
1) Maternal immune activation involves the maternal immune system. Immune activation is stimulated in the mother. It cannot be assumed that MIA is the same as causing immune activation in the fetus or in an infant receiving vaccines.
2) The fetus is at an earlier stage of development than an infant. It cannot be assumed that an infant can be similarly damaged by immune activation.
3) The immune activation experiments do not use vaccines. They use infections, lipopolysaccharide (LPS) or poly(I:C) to stimulate the immune system. It cannot be assumed that a vaccine will have the same effects as LPS or Poly(I:C). These substances can create very strong immune activation.
Vaccine advocates are mostly unaware of the growing literature on immune activation-mediated brain damage. But when they do become aware, they argue strongly that the science of immune activation is not relevant to vaccines for the reasons above.
This article shows these objections are contradicted by the scientific evidence. Vaccines given to an infant can cause brain damage by the same mechanisms proven to occur in maternal immune activation experiments.
Specifically, the science establishes the following:
1) Maternal immune activation (MIA) causes a cytokine surge in the fetal brain. It’s the cytokine surge that causes brain damage.
2) The brain can be damaged by immune activation postnatally, with no maternal involvement.
3) Brain development is a continuous process through gestation, birth, and infancy. The brain grows enormously during infancy. The developmental processes occurring after birth can be disrupted by immune activation.
4) Vaccine adverse reactions stimulate high cytokine production. Normal vaccine response does not cause brain damage. It’s the unusual vaccine adverse reactions that cause brain damage and autism.
In combination, these well-established facts prove that vaccines will cause brain damage, and autism specifically, in infants that suffer adverse vaccine reactions. Schizophrenia, depression, attention-deficient disorder and possibly other mental illness should also be expected.
1) The Fetal Brain During MIA
It is proven beyond any doubt that activation of the maternal immune system (during gestation) causes autism and other brain damage in the fetus. But what is the mechanism for this damage? Fortunately, there has been a lot of good research exploring this question.
The fetal brain is damaged by abnormal levels of cytokines in the brain. In some experiments, the cytokines come from the mother, and in others, the cytokines are produced by the fetal brain itself. The source of the cytokines seems to depend on the timing and number of immune activation events. In either case it’s the unusually high cytokine concentration in the developing brain that causes the damage.
Oskvig et al. (an NIH-funded study) describes a maternal immune activation study in which cytokine levels were measured in maternal serum (blood), amniotic fluid and fetal brains. The measurements were performed 4 and 24 hours after immune activation by lipopolysaccharide (LPS). Results are shown below. In this experiment, the fetal brain did not produce the cytokines (as determined by measurements of messenger-RNA expression). Instead, it appeared that cytokines produced by the mother traveled into the fetal brain.
Full paper (Oskvig): “Maternal Immune Activation by LPS Selectively Alters Specific Gene Expression Profiles”
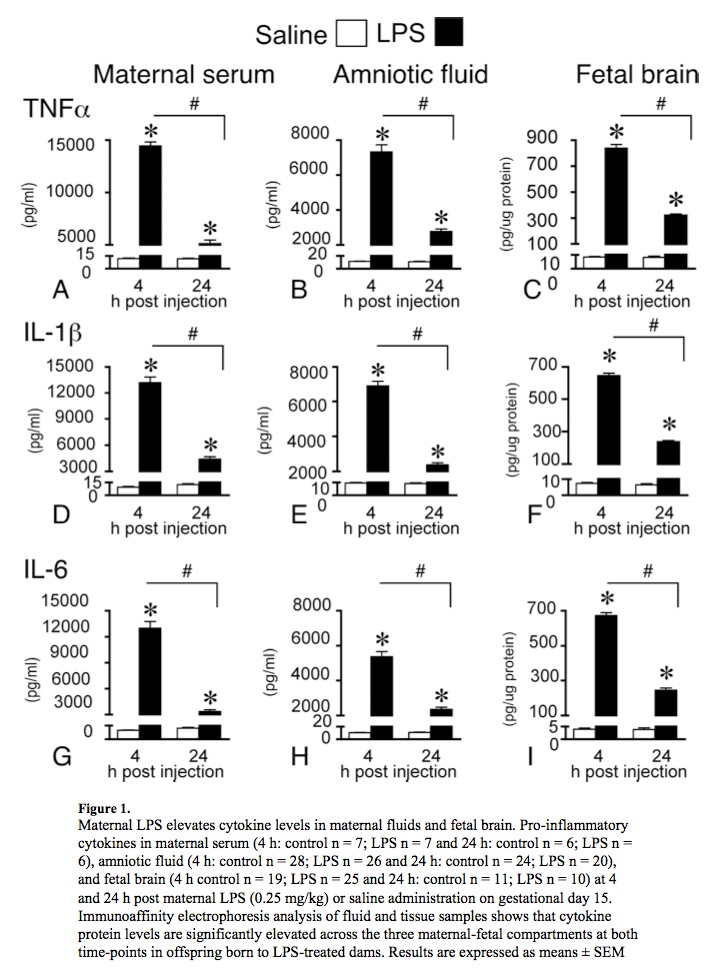
Above: Maternal immune activation (MIA) causes a cytokine surge in the fetal brain. In this experiment, cytokines appear to travel from the mother into the fetal brain. The animals in this experiment suffered brain damage and displayed autistic behavior. From Oskvig et al. 2012. LPS=lipopolysaccharide, an immune system activator.
Other MIA experiments using slightly different conditions (using either LPS or other immune activators) such as Ghiani et al. find that maternal immune activation does cause cytokine production within the fetal brain.
Ghiani et al. state:
Enhanced microglia activation, reactive astroglia and increased expression of pro-inflammatory cytokines were detected in the foetal brain . . . .
and
These data appear to indicate that an extended foetal response was induced, as maternal levels of reactive cytokines decreased rapidly, while foetal levels remained elevated well after exposure to LPS.
Full paper (Ghiani): “Early Effects of Lipopolysaccharide Induced inflammation on Foetal Brain Development in Rat”
This is clear evidence of immune activation in the fetal brain. In the experiment of Ghiani et al., the fetal brain is producing cytokines.
Why do Oskvig and Ghiani report contradictory results regarding cytokine production in the fetal brain? Oskvig describes two possibilities:
Microglia populate the embryonic brain during a surge of infiltration from external sources such as the yolk sac at about the time of the LPS injection in our study (Rigato et al., 2011). It is possible that they are not mature enough to respond to an immune challenge by making more cytokines of their own. Indeed, in preliminary studies, using CX3CR1-GFP heterozygous reporter mice, we find no evidence of microglial activation after maternal LPS injection given at the equivalent gestational age of day 13 (unpublished data).
A recent report by Ghiani et al. (2011) showed that following maternal LPS administered twice, on gestational days 15 and 16 in rats, there was evidence of activated microglia and astrocytes in fetal cortex two days later. In addition, both mRNA and protein levels of several inflammatory cytokines were elevated in the time window of 4-72 h following the LPS. A possible explanation for a fetal brain inflammatory response in that paradigm, not seen in our paradigm, may be attributable to the double injection procedure used in that study. Nevertheless, we emphasize that in our paradigm, despite the lack of evidence of gross changes in brain development and inflammation, the offspring displayed deficits in social and exploration behaviors as adults, indicating that the subtle changes occurring after a single maternal LPS injection are sufficient to cause altered behavior symptomatic of psychiatric disease.” (Emphasis added)
These details are important because vaccine advocates often assert that maternal immune activation is fundamentally different from immune activation during infancy (i.e., postnatally). However, the two conditions set forth above for immune activation in the brain (mature microglia, and multiple immune activation events) are both present in infants. Infants have mature microglia capable of producing cytokines, and vaccines are given in numerous exposures (typically at least 5 in the first 18 months: at birth, 2, 4, 6, 12 and 15 months).
The Ghiani et al. experiment may stimulate cytokine production in the fetal brain due to microglial priming, since two immune activation events were used. Microglial priming is a well known phenomenon in which microglia become increasingly sensitive and reactive to immune activation after repeated immune activation events. Each immune activation event makes the microglia more sensitive and produce more cytokines.
Both Oskvig and Ghiani found clear evidence of brain damage and pathological behavioral changes. The developing brain is damaged by cytokines whether they come from the mother, or are produced by the fetal brain (i.e., by microglia). In other words, the source of the cytokines is irrelevant. And the human postnatal brain can produce cytokines.
2) Look Mom, I Damaged It All By Myself!
Part 2 (on IL-6 and autism) described the important Wei et al. study in which mice exposed to IL-6 postnatally suffered autistic brain damage. This study was done by injecting a genetically engineered virus (containing the gene for IL-6) into the mouse brain. The gene caused chronic exposure of the brain to IL-6. The result was autistic behavior and physiological damage indicative of autism.
This result is important, and it is relevant to humans because the action of IL-6 is conserved across species and because the development of human and mouse brain are governed by the same basic principles. In fact, there are no known differences between humans and mice in the function of IL-6.
At birth, the human brain is more fully developed than the mouse brain, and therefore the human brain is arguably less sensitive to immune activation in the postnatal period.
This leads to an important question: can a more fully developed human brain suffer damage from immune activation? The answer is an unequivocal YES. There are numerous case reports of brain inflammation (“encephalitis”) causing autism in children at older ages. Some of the children recover, and some don’t. Here are 4 such case reports/case series and a review of the subject:
Case Series, DeLong 1981: This report describes 3 previously-normal children, ages 5(F), 7.5(M) and 11(F) that developed autism due to brain inflammation from an infectious illness. The authors state:
We report three cases in which striking autistic features developed in previously normal children in the course of an acute encephalopathic illness . . . .
and
These three children each demonstrated a full-blown autistic syndrome in the course of an acute encephalopathic illness . . . . The cases are presented as examples of an acquired and reversible autistic syndrome in childhood, affording some insight into the neurologic substrate of that syndrome.
and
. . . the behavioral syndrome was acquired at a clearly definable time, in the context of an acute encephalopathic illness . . . .
Full paper: “Acquired Reversible Autistic Syndrome in Acute Encephalopathic Illness in Children”
Case Report, Marques, 2013: This report describes a previously healthy 32-month old girl that suffered a viral infection of the nervous system. The child experienced “marked developmental regression, autistic features, persistent stereotypes and aphasia” (aphasia=loss of speech).
The authors described this case as “. . . encephalitis leading to developmental regression with autism spectrum disorder and correlating these 2 distinct entities.” (encephalitis = brain inflammation)
The authors rule out inborn metabolic disorders as a cause (this argument also applies to the other case reports):
Our patient was previously healthy, with adequate psychomotor development until this infectious episode. Additionally, newborn screening and metabolic tests performed on the admission were negative (normal values for ammonia and lactate) and MRI images do not suggest metabolic disorders. Finally and most importantly, she had a favorable outcome with improvement in all skills and development quotient enhancement. These features do not support an inborn error of metabolism, which are characteristically progressive and lead to severe mental retardation.
Full Paper: “Autism Spectrum Disorder Secondary to Enterovirus Encephalitis”
Case Report, Ghaziuddin, 2002: This report describes a previously healthy 11-year-old boy that developed illness with fever, seizure, and brain inflammation. In the following months, the child developed autism and never fully recovered. The authors state
On the DSM-IV symptom checklist for autistic disorder, he met all the criteria for autism except the onset criterion because he did not have a history of any symptoms before three years of age.
and the authors state that this case “provides further evidence that autistic symptoms can sometimes emerge after the age of three years following an external event such as an infection.
Full Paper: “Autistic Symptoms Following Herpes Encephalitis”
Case Report: Gillberg, 1986: This report describes a previously healthy 14-year-old girl that developed illness with fever, seizure, and brain inflammation. In the following 70 days, she developed all the symptoms of autism, including echolalia, loss of speech, hand-flapping and other self-stimulating behavior. She never recovered. Gillberg states
I will describe the case of a 14-year-old-girl who developed all the classical symptoms of autism over a 70-day period after the onset of convulsions in herpes simplex encephalitis (presumptive diagnosis on the basis of clinical, neurochemical, and neurophysiological tests). Only the age criterion is not met, since she was totally psychiatrically healthy up to her 14th year of life.
and
Severe autistic features — including gaze aversion, abnormal reactions to sound, delayed and immediate echolalia after initial muteness, typical hand-flapping stereotypies, and the acquisition of a set of routines and insistence on sameness that make everyday life totally restricted — developed over a 3-month period after the onset of encephalitis and still remain.
Full Paper: “Onset at Age 14 of a Typical Autistic Syndrome: A Case Report”
Review, Libbey, 2005: This review describes the prior case reports of infection-induced brain inflammation causing autism. At the time this review was written in 2005, immune activation was increasingly suspected as a cause of autism. The authors make the obvious connection between cytokines and infection:
An acute infection could lead to transient levels of cytokines without viral persistence, or infection could instigate an autoimmune process resulting in chronically elevated cytokine production . . . . Depending on the location of immune activation, cytokines could be produced directly in the brain or gain access to the CNS [central nervous system] by crossing an immature blood-brain barrier (BBB).
Full Paper: “Autistic Disorder and Viral Infections”
Ten years of intense research since the Libbey review paper has firmly established that brain inflammation and cytokines (specifically IL-6 and IL-17a) cause autism. It is no longer debatable whether cytokines cause autism.
The above case reports demonstrate that the human brain can be damaged by immune activation and cytokine surges in older children. The human brain remains vulnerable for many years, though the evidence suggests that sensitivity declines with age. The argument by vaccine advocates that the human brain is vulnerable to immune activation only while in the womb — i.e., that maternal involvement is necessary — is contradicted by the above case reports. The human brain can suffer autistic damage without any maternal involvement whatsoever.
In order for vaccine advocates to assert that the infant brain is not vulnerable to immune activation, they must argue that the age range of about 0-3 years is especially privileged and uniquely resistant to immune activation. This idea is preposterous and is contradicted by even more evidence in argument #3 below on brain development processes.
More Evidence of Postnatal Damage
There is even more evidence from animal experiments that postnatal immune activation causes autism brain damage. For example, a recent (2015) maternal immune activation study by Coiro et al. demonstrated that ongoing immune activation in the postnatal period accounted for most or all of the damage. This was demonstrated by giving MIA-exposed infant mice an anti-inflammatory drug (ibudilast) postnatally. Ibudilast is known to reduce inflammation in the brain. The ibudilast treatment prevented autism damage. The authors state
. . . we found that postnatal treatment with an anti-inflammatory drug can prevent the dendritic spine loss as well as the increased marble burying in MIA [MIA=maternal immune activation] offspring. We suggest that an altered inflammatory state in the developing brain of MIA offspring affects synaptic development and behavior.
and
These data suggest that increased inflammatory state during early postnatal weeks is responsible for altered synaptic connectivity and impaired behavior and that early anti-inflammatory treatment can have an ameliorating effect on both synapses and behavior. (Emphases added)
Full Paper (Coiro): “Impaired Synaptic Development in a Maternal Immune Activation Mouse Model of Neurodevelopmental Disorders”
Results illustrating the protective effect of ibudilast are shown below.
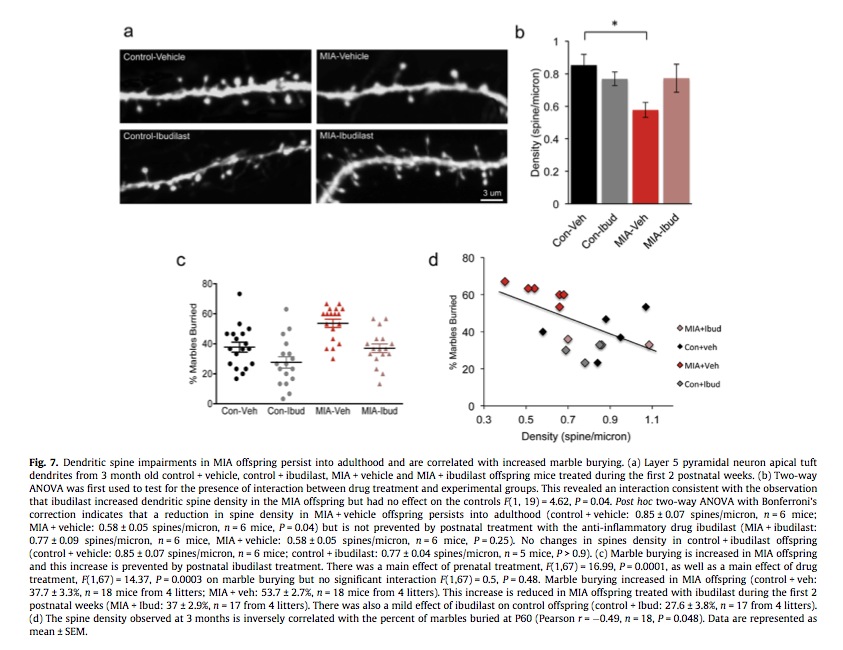
Above: The anti-inflammatory drug ibudilast given postnatally prevented the behavioral changes and synapse damage caused by maternal immune activation (MIA). This implies that the brain is damaged by inflammation in the postnatal period. From Coiro et al. 2015.
Con=control group (not exposed to MIA)
Veh=vehicle (vegetable oil carrier for dissolving ibudilast)
Ibud=ibudilast (an antiinflammatory drug that reduces inflammation in the brain)
Studies Of Postnatal Immune Activation
There are several studies on the brain effects of immune activation in the postnatal period. This research is not on autism specifically, but instead looks at seizure disorders and synaptic development (and other outcomes). Autism and seizure disorders are strongly associated. About 30% of autistics have a seizure disorder.
Note that postnatal days 10 and 15 in rats corresponds to birth and age 1-2 years in humans, respectively.
Li et al 2015: This study measured the effects of two different vaccines (BCG and Hepatitis B) given postnatally on brain development in mice. The Hepatitis B vaccine caused adverse effects on brain development, and the BCG vaccine caused beneficial effects. The opposite effects are attributed to the opposite types of immune activation stimulated by the vaccines. [Editor’s note: This may help to explain why countries that administer BCG vaccines at birth do not tend to have comparable autism rates to countries that administer hepatitis B vaccines at birth.] This study is described in detail here:
Harre et al 2008: This study observed behavioral deficits in rats from immune activation on postnatal days 5 and 30. This study states “Behavioural deficits were observed specifically in the P5 and P30 LPS-treated groups in the water maze probe trial and fear conditioning tests . . . . ” and
Thus, a single bout of inflammation during development can program specific and persistent differences in NR mRNA subunit expression in the hippocampus, which could be associated with behavioural and cognitive deficits in adulthood.
Galic et al 2008: This study observed increased seizure susceptibility in rats from immune activation on postnatal days 7 and 14. This study states “These persistent increases in seizure susceptibility occurred only when LPS was given during a critical postnatal period (P7 and P14) and not before (P1) or after (P20).” and “These novel results indicate that a single LPS injection during a critical postnatal period causes a long-lasting increase in seizure susceptibility that is strongly dependent on TNFα.” Note: TNFa is an inflammatory cytokine induced by immune activation.
Paper: “Postnatal Inflammation Increases Seizure Susceptibilty in Adult Rats”
Chen et al 2013: This study observed increased seizure susceptibility and impaired brain brain development in rats from immune activation on postnatal day 14. This study states
These data indicate that systemic inflammation . . . during the critical stages of childhood development may affect the immune system into adulthood by increasing susceptibility to seizures and the vulnerability of neurons to seizure damage and affecting the synaptic plasticity in the hippocampus.
These studies clearly demonstrate brain damage in rats at postnatal days that correspond to human ages when infant vaccines are recommended (about age 1-2 years).
3) Human Brain Development During Vaccination
While there are many similarities between human and mouse brain development, there are important differences in timing. Brain development processes are very similar in humans and mice, but they occur in different sequences and at different times relative to other developmental events. For example, the human brain is more fully developed at birth than the mouse or rat brain; 7-day-old rats are considered to be comparable in brain development to a newborn human brain. These issues are complicated and so will not be covered here. See this paper by Semple et al. if you are interested (WARNING: may cause drowsiness):
Full paper (Semple): “Brain Development in Rodents and Humans: Identifying Benchmarks of Maturation and Vulnerability to Injury across Species”
A logical way to try to determine if the human brain is sensitive to immune activation postnatally is to consider the specific development processes happening at this time. Are the development processes occurring in the postnatal human brain sensitive to immune activation and disruption by cytokines? Scientific evidence shows that some developmental processes occurring in the human brain postnatally — when vaccines are administered — are indeed disrupted by immune activation and cytokines.
Fundamental brain development processes include:
Neurogenesis: The birth of new neurons. Neurons carry information in the brain. Most neurogenesis occurs early in gestation.
Gliogenesis: The birth of new glial cells. Glia include immune system cells of the brain (microglia).
Synaptogenesis/synapse formation: The formation of synapse connections between neurons. Synaptogenesis is intense for the first few years after birth.
Synapse pruning: The elimination of unneeded synapses. Synapses are greatly overproduced early in life and then slowly eliminated throughout childhood and adolescence.
Myelination: The formation of the myelin sheath around neurons. Myelination begins in late gestation and continues for years after birth.
Migration: The movement of neurons and other brain cells toward their final positions. Occurs during gestation and after birth.
Neural Adhesion: The formation of chemical-mechanical attachments between neurons and glial cells. These attachments hold the cells in place. Adhesion occurs after neurons migrate to their proper locations, which is logically expected.
Wei et al. demonstrated that cultured infant mouse neurons suffer defects in adhesion, migration and synaptic formation when exposed to IL-6. Specifically, synapse formation is altered to produce an excess of excitatory synapses, a deficit of inhibitory synapses and changes indicative of autism. Human autistics show many signs of excessive neuronal excitation (lack of pre-pulse inhibition, sensitivity to lights and sounds, stimming etc.). *
Wei et al. state
These findings suggest that the elevated IL-6 in the autistic brain could cause an imbalance of neuronal circuits through its effects on neural cell adhesion/migration and synapse formation, and contribute to the development of autism. (Emphasis added)
Full Paper (Wei): “IL-6 is Increased in the Cerebellum of Autistic Brain and Alters Neural Cell Adhesion, Migration and Synaptic Formation”
Are adhesion, migration, or synapse formation occurring when vaccines are administered? YES. Synapse formation is intense during the first two years of life. Not as much is known about the timing of adhesion and migration in the human brain.
The sequences and approximate time periods for brain development processes in human brain are mostly known. Below is a chart illustrating the timing of these processes and vaccination. Unfortunately, the chart does not show adhesion or migration. Migration is mostly complete before birth, and continues at a much slower rate postnatally. I have been unable to find reliable literature on the timing of neural adhesion in humans. It appears that the time course of adhesion in human brain development is not yet known. Since adhesion occurs after migration (and in association with synapse formation) and migration continues postnatally, adhesion must occur postnatally as well.
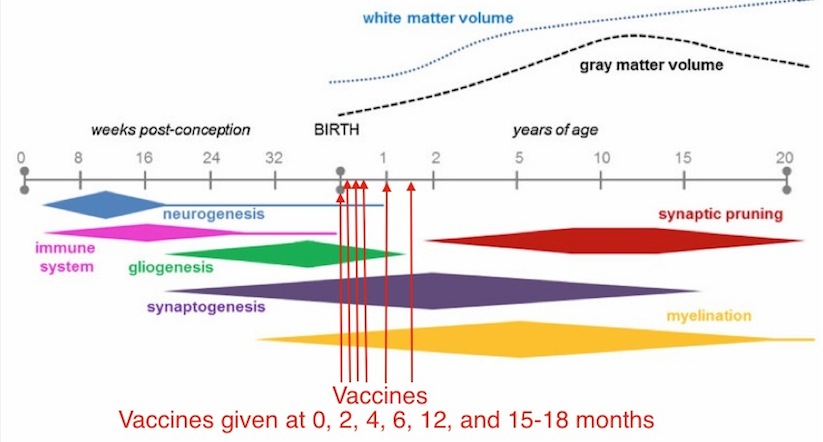
Above: Vaccination occurs during intense synapse formation in the human brain. Synapse formation is disrupted by immune activation and cytokines, which are stimulated by vaccine adverse reactions. Image adapted from Semple et al.
The human brain experiences an intense burst of synapse formation immediately after birth which continues for over a year. For example, the Semple et al. paper states
While the proliferation of synapses begins around 20 weeks of gestation, density increases rapidly after birth, particularly within the early postnatal months, to reach a level approximately 50% higher than that seen in adults by 2 years of age. (Emphasis added)
The literature on synaptogenesis describes synaptic formation as “exuberant.” For example, a 2010 review on brain development by Stiles et al. states “ . . . studies of both monkeys and humans have documented widespread exuberant production of connections throughout all brain regions in the early postnatal period.”
Coiro et al. supports the idea that inflammatory cytokines during synaptogenesis are a cause of the damage:
Our data suggest therefore that reduction in inflammatory cytokines during the period of synaptogenesis in MIA offspring has long lasting effects indicating a therapeutic potential of ibudilast in treating MIA induced neurodevelopmental disorders. (Emphasis added)
Full Paper (Stiles): “The Basics of Brain Development”
The first high-quality measurements of synapse formation in human brain were by Huttenlocher et al. in 1997. The most important data from this study is provided below. It clearly shows a dramatic increase in synapse density in the first year of life. Consider that the brain volume/weight is also increasing during this time (from 400g to 900g in the first year), so the increase in the total number of synapses is even greater than the increase in density. Synapse formation in the first year of life is truly “exuberant.”
Above: Synapse formation in humans is intense during the first year after birth, when vaccines are administered. Synapse formation is disrupted by immune activation and cytokines, which are stimulated by vaccine adverse reactions. Image adapted from Huttenlocher et al.
Paper (Huttenlocher): “Regional Differences in Synaptogenesis in Human Cerebral Cortex”
Vaccines are administered during intense synapse formation. If vaccines induce IL-6 during this time, the result will be an excess of excitatory synapses, deficit of inhibitory synapses, and disturbances of neuronal migration and adhesion (to the extent they are occurring). The result is autism brain damage and behavior.
Conclusion
The evidence is consistent and strong. Sensitive brain development processes continue after birth. The deloping infant brain is damaged by immune activation, just like the fetal brain. The developing human brain can be damaged by immune activation in the postnatal period, when vaccines are administered.
Part 5 in this series shows that vaccine adverse reactions stimulate IL-6 in the brain: Part 5: Vaccine and Immune Activation
_____________________________________________________________________
*Excitation/Inhibition Imbalance in Human Autistics
It is generally accepted that human autistics have an excitatory/inhibitory imbalance. Until recently, this was based on strong but circumstantial evidence. Ecitatory/inhibitory imbalance is consistent with what is known about autistic behavior and animal models, but there has not been direct experimental evidence for it, until now (December 2015). Robertson et al. report experimental evidence for excitatory/inhibitory imbalance in autistic subjects. In this experiment, two different images are presented to a subject, one to each eye. In this situation, the left (L) and right (R) visual circuits compete for perceptual dominance. The subject’s attention switches back and forth between the images. This “perceptual switching” requires inhibition of the active visual circuit (the L or R circuit). Since autistics are less able to suppress neuron activity, they report much less frequent perceptual switching. Also, autistics are less able to consciously control perceptual switching. Normal control subjects are able to switch their perception much more rapidly and easily. Also, autistics are less able to fully inhibit perception of an image. Robertson et al. state
An imbalance between excitatory/inhibitory neurotransmission has been posited as a central characteristic of the neurobiology of autism . . . .
and
We demonstrate a robust, replicated autistic deficit in binocular rivalry, a basic visual function that is thought to rely on the balance of excitation/inhibition in visual cortex . . . .
and
These results suggest a disruption in inhibitory signaling in the autistic brain . . . .
Full Paper (Robertson): “Reduced GABAergic Action in the Autistic Brain”
Click here for a list of papers in this post.
Click here to see other posts in this series.










I know this site predominately hesitant to promoting vaccines(as I am myself/very leary doing any). I’m wondering is there an age where it is safer to start vaccinating? Also if you do one at a time, does that help? Just curious due to school requirements. This is so tough bc hard to get around them at times but I definitely agree they contribute to autoimmune/neuro. Thanks in advance.
Ah, Nicole, you DO have to consider all the conditions you face. As far as brains are concerned, it is safER after five when brain development has slowed, but it is still going to have negative consequences for at least a proportion of the population. There are a lot of genetic factors that affect the immune response, many of which affect methylation and sulfation, two ways the body clears itself of toxins. People who have challenges in these areas are much more likely to react badly to the immune activation that vaccines trigger. One at a time is also likely to help, as there would be fewer synergistic effects. Most serious injuries seem to occur after children received multiple vaccines at once, but a number of our vaccines today are themselves multiple vaccines.
Yes! I agree 100%. My 9 year old son I believe suffered damage, he had numerous high fevers months to years after the vaccines. He now has ADHD and it’s tough to deal with. I still feel this may have been caused/accelerated due to the vaccines. I stopped vaccinating midway with my 2nd son bc I noticed neurological changes in him. Thank God I followed my gut. My 3rd child/girl has had very few bc she had a high fever after 1 and I was done, she’s now 4 but I’m having to take her from a school she loves because she’s not vaccinated:( I wanted to wait even longer to do anymore if I even decide to do them but this makes me feel better knowing I made the right choice not doing them. The damage is done once they’ve received the shot I believe. I do have an mthfr mutation so I wonder if they may have and can that cause reactions? Also would like more info to detoxing or supplementing with vitamins that could help. Thanks so much for answering me, it makes me feel I made the right decision taking my daughter from the school. I will wait until she’s a little older if we choose to do anymore. Such tough decisions is parents face. Thanks for all you do.
The school nurse also told me that a reaction would happen right away. I believe this is not true in all the info I’ve read. I didn’t even bother debating with her.
MTHFR makes it harder to detoxify, so yes it is related to more intense vaccine reactions. There are lots of places on the web that discuss the various supplements recommended for specific genetic situations, including Amy Yasko, http://www.mthfrsupport.com.au/, mthfr.net. One of the best means for detox is the AMD IonCleanse footbath. A number of TMR moms did a study with it, and many of their children experienced huge gains as a result, including older children that began talking.
Where do you live? In the United States there are only three states that do not accept religious exemptions, and they are very easy to obtain in most states. You can check out your state’s regulation at NVIC.org.
And you’re welcome. 🙂
Professor TMR - if you are there
the best article I’ve seen on AoA in a while :
http://www.ageofautism.com/2016/10/who-is-telling-flu-vaccine-truth-the-cdc-or-vaccine-makers.html
Was the MRI given to the 32 month old female patient in Case Report, Marques, 2013: with or without Gadolinium Contrast?! We are finding HIGH amounts of this drug in many patients post MRI with Gadolinium contrast by nearly seven years post exposure, in patients with NO KNOWN KIDNEY DISEASE!
Please see the site:
http://www.gadoliniumtoxicity.com
for up to date research information. I am trying to save those children with Autism from the the same fate that is befalling myself; I am 57 with no known kidney disease yet have systemic fibrosis in many areas of my body, NOT just dermal fibrosis or focal dermal fibrosis with increased mucin. I have Fibrosis of the bowel, yellowing in the left scleral body (whites of the eye) hardened woody skin, eleven dermal biopsies, etc.
Being told I will NEEVR PROVE THAT GADOLINIUM DID THIS TO ME, is the last straw! No one should be LIED to AFTER they are poisoned! I am also affected via vaccines, receiving Rubella at my son’s birth in 1989, bleeding for 17 months postpartum, having several D&C’s and later into my forties, two colorectal surgeries to repair tissue damaged because my insides are “hardening/scarring/fibrosing” Still later when I did not realize the Rubella was doing this things to me, I allowed myself to be vaccinated against Tetanus ( I have NTO stopped spasming since, so severely the left side of my throat has scarred, shortened in length!) ad Pneumonia, when I never had pneumonia as one of seven children, yet since that fatefuk injection fo infection, I have succumbed to Klebsiella Oxytoca Pneumonia at least a dozen times, being hospitalized at least 5 times, three of them within a one year timespan in 2012. In 2014, while having my cancerous thyroid removed along with the left elongated and calcified styloid bone removed, I had Pneumonia as well (per required chest xray in an over 50 year old surgical patient!) and THEY STILL WENT AHEAD WITH THE SURGERY TO REMOVE THE THYROID AND STYLOID!
I want to let each and every porent and caregiver of children, please go to the http://www.gadoliniumtoxicity.com site above and check out every single piece of research posted there; if your child HAS had an MRI with Gadolinium contrast look me up on Facebook or Twitter, you will need further help for what is about to happen with your kids, LOVE THEM, HUG THEM, NEVER VACCINATE THEM!
IT is POISON as is GADOLINIUM CONTRAST!
PS: This article is a wonderful piece of research!
Absolutely love these graphs !
And they tie beautifully with the Autism ONE talk given by Brian Hooker in 2014
Wonderful article .